
GROWTH AND DEVELOPMENT
Growth is a fundamental characteristic of living organisms.
OR
Growth is defined as an irreversible increase in dry mass of living material.
OR
Growth is a permanent or irreversible increase in dry mass of protoplasm due to synthesis of proteins.
Dry mass is mass without water.
Why dry mass?
By specifying dry mass, we can ignore the short-term fluctuations in the water content of the cells, for instance when plant cells take in water by osmosis. The reverse process can take place when cells lose water.
Other definitions of growth:
- Growth as increase in size. This definition is inadequate; some organisms, e.g., plants, can increase in size as they take in water by osmosis, but this process may be reversible when they lose water.
- Growth as increase in cell number.
This definition is inadequate when the zygote divides repeatedly to form a ball of cells, the early embryo; there is an increase in cell number without increase in size of daughter cells.
In some cases, you can increase in size without increase in cell number, e.g., in the region of elongation behind the root and shoot tips.
- Growth as increase in number of individuals (population of single-celled organisms, e.g., microorganisms such as bacteria).
Development: The process of development is so closely linked with growth that the phrase “growth and development” is commonly used to describe processes normally thought of as growth. Development refers to an increase in complexity due to differentiation of tissues and organs (improvement in the functions of the body).
Growth can be regarded as change in quantity while development is the change in quality.
Factors that affect growth
There are both external and internal factors that control growth.
| External factors | Internal factors |
|---|---|
| Food/nutrients | Genes |
| Diseases | Hormones |
| Temperature | Enzymes |
| Oxygen | |
| Light | |
| Space | |
| Toxins | |
| Soil | |
| Carbon dioxide (CO2) |
Patterns of growth
- Positive and negative growth
Positive growth occurs when synthesis of materials (anabolism) exceeds breakdown of materials (catabolism).
An example of positive growth in plants is the production of seedlings, which involves increase in cell number, cell size, fresh mass, length, volume, and complexity of form as the seedling starts to photosynthesize and make its own food.
Negative growth occurs when catabolism exceeds anabolism. For example, a decrease in dry mass of germinating seeds.
- Allometric and isometric growth
Allometric growth occurs when organs grow at different rates. This produces a change in size of the organism accompanied by a change in shape.
This pattern of growth is characteristic of animals. In almost all animals, the last organs to develop and differentiate are the reproductive organs. In humans, the heart, brain, and gonads all have different growth rates.
Isometric growth occurs when organs grow at the same rate. In this case, change in size of the organism is not accompanied by a change in shape or external form.
This type of growth pattern is seen in fish and certain insects such as locusts.
- Limited and unlimited growth
Limited growth (definite/determinate) is the type of growth which ceases when an organism matures and reaches reproductive age. For example, growth in annual plants.

Unlimited growth occurs throughout the life of an organism. This growth mostly occurs in perennial plants and is characterized by a series of sigmoid curves.

Measuring growth
Growth can be measured using different parameters, such as length/height, mass (dry/fresh), weight, volume, and area.
Growth curves
Growth curves are graphs obtained when data from different parameters of measuring growth are plotted against time.
The curves show the overall growth pattern and extent of growth. It has been found that growth patterns in many organisms tend to be similar regardless of the parameter used. In many cases, if an increase in a measurable parameter is plotted against time, an S-shaped growth curve is obtained. The shape of these curves is described as sigmoid, meaning S-shaped. The term sigmoid is derived from the Greek word sigma meaning letter S. A sigmoid curve is divided into four parts or phases.
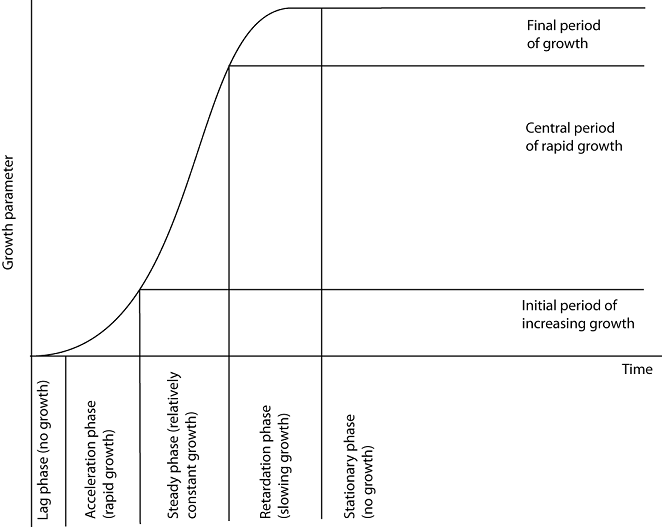
- Lag phase
This is the initial phase during which little growth occurs (sometimes a slight decrease). In flowering plants, this phase shows a slight decrease in growth due to loss of dry mass during seed germination.
In microorganisms, some may die at the time of inoculation to the culture medium, showing a decrease in number. During this phase, the population of microorganisms grows slowly because they may have been dormant and require time before their metabolism becomes efficient. Another reason for little growth may be adjustment to the new environment.
- The log phase or exponential phase
This is the period of rapid growth during which growth proceeds exponentially. The rate of growth is at maximum and proportional to the amount of material or number of cells or organisms present. In microorganisms, this phase occurs when there is no limiting growth factor; nutrients, oxygen, and space are plentiful. In flowering plants, it is the period when green follicles increase in amount.
- Stationary phase
This marks the period where overall growth ceases. The parameters remain constant. In microorganisms, the number stabilizes; the number dying equals the number formed.
- Decelerating phase (decline phase)
This is the period when growth becomes limited due to internal and external factors or their interaction. In many mammals including humans, this marks negative growth and senescence associated with aging.
In microorganisms grown in confined environments, this is when the carrying capacity declines and cannot support high density. Nutrients decrease and waste products increase, leading to death due to starvation, oxygen shortage, or toxic waste.
Diagram showing a sigmoid curve
- Growth in arthropods
Growth in arthropods occurs in a series of stages called instars. These stages show sudden changes in weight or length. This pattern is known as intermittent or discontinuous growth. Each stage is called an instar.
Reasons for intermittent growth
All arthropods have an exoskeleton made of hard chitinous cuticle, which prevents overall growth of the body. The exoskeleton is shed periodically in a process called moulting or ecdysis to allow growth.
The new cuticle underneath is soft enough to allow growth. It later hardens, making growth impossible until the next moult. Thus, growth occurs in spurts interrupted by moults.
Diagram showing growth in arthropods
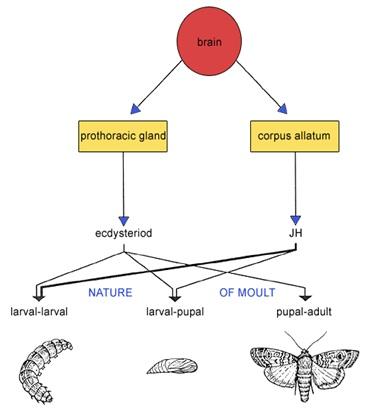
Hormonal control of ecdysis or moulting in insects
Moulting or ecdysis is controlled by a moulting hormone (mH) or ecdysone, released in response to a specific stimulus. The hormone is secreted by the thoracic gland, stimulated by hormones from neurosecretory cells in the brain.
Moulting hormone is a steroid that causes shedding of the cuticle and growth of the insect. Growth is accompanied by a series of moults. The hormone causes secretion of moulting fluid beneath the cuticle, dissolving the soft inner part while the epidermis secretes a new soft cuticle.

Growth in insects
Growth in insects involves changes in body form through stages in their life cycle, from young to adult. These changes are called metamorphosis.
Metamorphosis also occurs in other groups such as amphibians, mollusks, crustaceans, trematodes, cestodes, and echinoderms. It refers to rapid changes during transition from larva to adult.
Metamorphosis in insects
There are two types of metamorphosis:
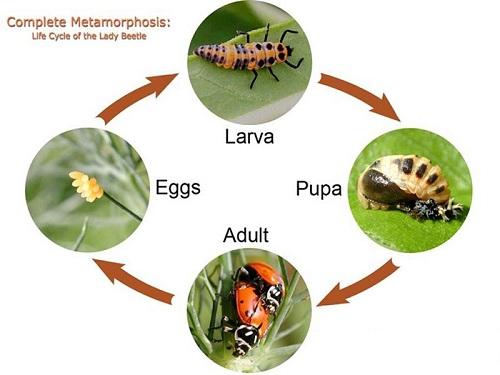
- Complete metamorphosis
In this type, the insect life cycle passes through four stages: egg, larva, pupa, and adult.
Insects with this type are called holometabolous, e.g., butterflies, houseflies, moths, and beetles.
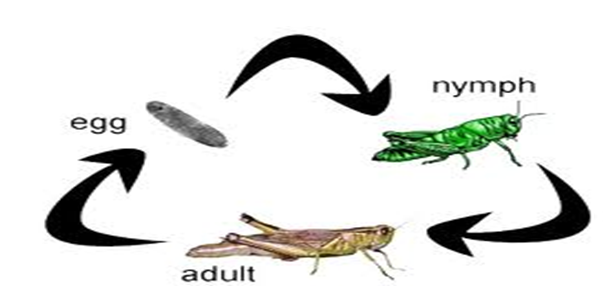
- Incomplete metamorphosis (hemimetabolous)
In this type, the insect passes through three stages where the young resembles the adult: egg, nymph, and adult.
Insects with this type are called hemimetabolous, e.g., cockroaches, grasshoppers, and mosquitoes.
Hormonal control of metamorphosis in insects
Successive moults lead insects to acquire adult features either suddenly or gradually. Metamorphosis is controlled by two hormones:
- The moulting hormone.
- Juvenile hormone.
Metamorphosis is suppressed by juvenile hormone secreted by the corpus allatum gland in the brain.
In the presence of juvenile hormone, epidermal cells under moulting hormone produce juvenile cuticle, resulting in nymph or larva. Juvenile hormone inhibits metamorphosis by suppressing genes for adult structures.
At metamorphosis, corpus allatum stops secreting juvenile hormone, and moulting hormone alone causes adult cuticle formation.
- Moulting hormone is required for shedding the cuticle.
- Moulting hormone with juvenile hormone causes larval cuticle production.
- Moulting hormone alone causes adult cuticle production.
The process of growth
Growth of multicellular organisms can be divided into three phases:
- Cell division – increase in cell number by mitotic division.
- Cell expansion – irreversible increase in cell size due to water uptake and synthesis of materials.
- Cell differentiation – specialization of cells.
a) Cell Division
Cells form from pre-existing cells by cell division, which involves division of cytoplasm into two daughter cells. The daughter cells share duplicated organelles.
Two major events in new cell formation:
- Division of the nucleus (karyokinesis).
- Division of the cytoplasm (cytokinesis).
Nuclear division
Two types of nuclear division:
1. Mitosis – nucleus divides to produce two daughter nuclei with identical chromosome sets to the parent. Maintains diploid chromosome number. Occurs in somatic cells, forming body cells.
2. Meiosis – nucleus divides to produce four daughter nuclei each with half the chromosome number. Known as reduction division, reducing diploid (2n) to haploid (n). Occurs in gonads, forming sex cells.
Note: Nuclear division involves chromosome distribution, which carry hereditary information.
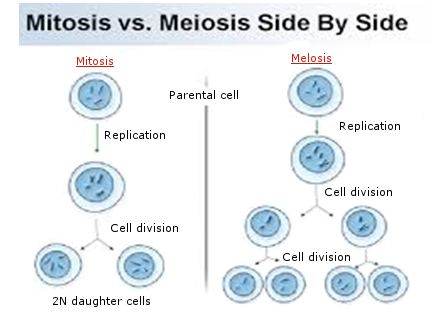
The cell cycle
The cell cycle is the sequence of events between formation of a cell and its division. It has three main stages:
- Interphase
Interphase is a period of intense synthesis and growth, producing materials for growth and other functions. It is divided into:
i. G1 (Gap one) or first growth phase
- Production of mitochondria, chloroplasts (plants), lysosomes, ER, Golgi complex, vacuoles.
- Formation of structural and functional proteins.
- Production of RNAs.
- Ribosome synthesis.
- High metabolic rate.
ii. S (synthetic phase)
- DNA synthesis.
- Production of histones covering DNA strands.
- Chromosomes become two chromatids.
iii. G2 (Gap two) second growth phase
- Centriole replication.
- Mitotic spindle formation begins.
- Energy stores increase.
- Intensive RNA and protein synthesis.
- Mitosis (M) – nuclear division separating chromatids into daughter cells.
- Cell division – division of cytoplasm into two daughter cells.
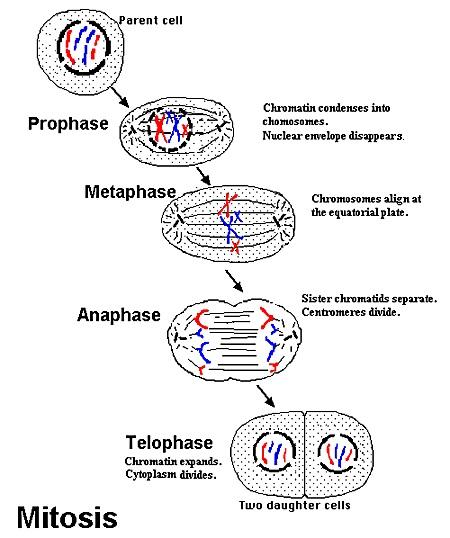
Process of mitosis
Mitosis is continuous with four active stages: prophase, metaphase, anaphase, and telophase. Interphase occurs between divisions. Stages in animals:
1. Prophase
- Chromosomes appear as chromatids joined by centromere.
- Nuclear membrane disintegrates.
- Nucleoli disappear.
- Centrioles move to opposite poles.
- Microtubules radiate from centrioles forming asters.
2. Metaphase
- Spindle fibres form.
- Chromatids attach to spindle at centromere.
- Nuclear membrane and nucleoli disappear.
- Chromosomes line up at spindle equator.
3. Anaphase
Rapid stage:
- Centromere splits into two.
- Daughter centromeres pulled to opposite poles by spindle fibres.
- Separated chromatids (now chromosomes) follow centromeres.
4. Telophase
- Chromosomes reach poles.
- Chromosomes uncoil, lengthen, form chromatin.
- Spindle fibres disintegrate; centrioles replicate.
- Nuclear membrane and nucleoli reappear.
- Leads to cytokinesis.
- Cell division (cytokinesis)
Cell division divides cytoplasm into two daughter cells. Organelles distribute between cells. Cytoplasmic division differs in animals and plants.
Cytokinesis in animals
Cell membrane invaginates at spindle equator, membranes meet at center, dividing cell into two daughter cells.
Cytokinesis in plants
Spindle fibres form a cell plate across equator, which develops into new cell wall dividing the cell.
Difference between mitosis in plants and animals
| Plants | Animals | |
|---|---|---|
| 1 | No centriole present. | Centrioles present. |
| 2 | No aster forms. | Aster forms. |
| 3 | Cell plate forms. | No cell plate forms. |
| 4 | No furrowing of cytoplasm at cytokinesis. | Furrowing of cytoplasm at cytokinesis. |
| 5 | Occurs mainly at meristems. | Occurs in tissues throughout the body. |
Significance of mitosis
- Growth and development
Mitosis increases body cell number, enabling growth.
Body repair replaces worn-out cells by new cells formed by mitosis.
New cells can differentiate to form complex structures.
- Genetic stability
Mitosis produces nuclei with the same chromosome number and hereditary information as parent cells, ensuring genetic identity and stability.
- Asexual reproduction
Many species propagate asexually by mitotic cell division.
- Regeneration
Some organisms replace lost parts (e.g., crustacean legs) through mitosis.
Seed germination
Germination is the onset of embryo growth in seeds or transformation of a seed into a seedling.
Environmental conditions needed for germination
1. Water
Water activates biochemical reactions in germination, many of which occur in aqueous solution. It also hydrolyzes stored food. Water enters seeds through the micropyle and seed coat by imbibition.
2. Temperature
Seeds require minimum or optimum temperatures (5 to 40°C) for germination. Temperature influences enzyme-controlled reaction rates.
3. Oxygen
Oxygen is needed for aerobic respiration, releasing energy by oxidizing food. Anaerobic respiration may supplement in some cases.
Physiology of seed germination
Seeds store carbohydrates, lipids, proteins, mineral salts, and vitamins. Major reserves are lipids and carbohydrates; starch is common in grasses and cereals, proteins in legumes.
Food is stored in endosperm or cotyledons (endospermic and non-endospermic seeds). In beans, cotyledons store food used for energy and building tissue before photosynthesis begins.
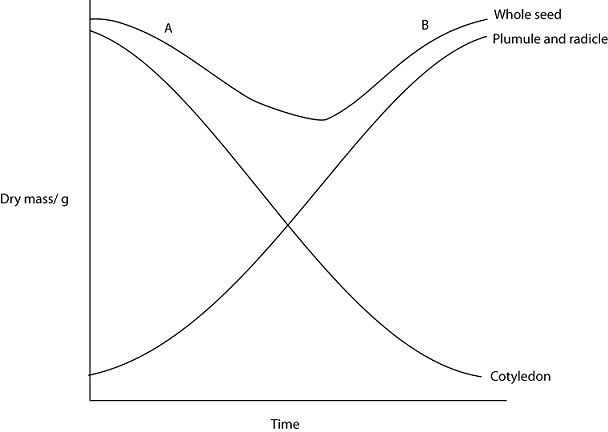
- Water uptake by imbibition and osmosis hydrates food reserves, activating respiration enzymes and synthesizing other enzymes.
- Digestion (hydrolysis) of food reserves; soluble products translocate to embryo growth regions.
- Oxidation of substrates (usually sugar) releases energy for storage tissue and embryo growth.
Respiration causes dry mass loss due to sugar loss; water is excluded in dry mass measurement. High respiration rates occur in storage tissue and embryo due to intense metabolism. Dry mass loss continues until seedling produces green leaves and photosynthesizes.
Types of germination
Two types based on cotyledon position relative to soil:
- Epigeal germination
Cotyledons are carried above ground. In dicots, the hypocotyl elongates, carrying cotyledons above soil. The hypocotyl remains hooked to protect the plumule tip during soil penetration and strengthens upon sunlight exposure.

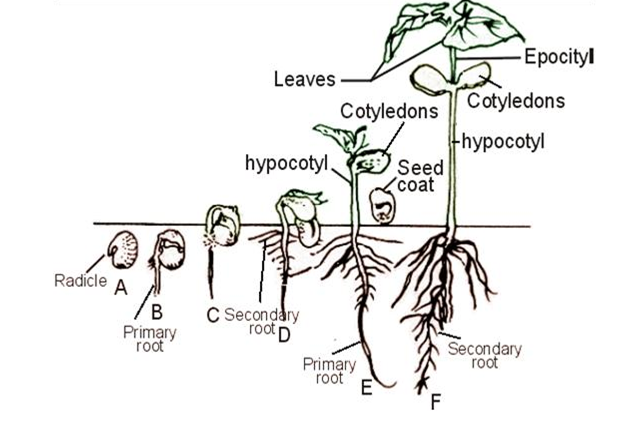
- Hypogeal germination
Cotyledons remain below ground. The epicotyl elongates, keeping cotyledons underground. In dicots, the epicotyl is hooked to protect the plumule tip and straightens upon sunlight exposure. In monocots like grasses, the plumule is protected by a coleoptile sheath; the first leaf grows through and unrolls in response to light.
Growth of the embryo
The first sign is emergence of the radicle (embryonic root), which grows downward and anchors the seed. The radicle is positively geotropic.
Next is the plumule, which grows upward and is positively phototropic.
Primary and secondary growth in flowering plants
Except for the young embryo, plant growth is confined to meristems—specific regions such as root and shoot tips.
Meristems
A meristem is a group of cells retaining the ability to divide by mitosis, producing daughter cells that grow and form the plant body. Cells losing this ability form permanent tissue.
Meristems, also called initials, are classified by location:
- Apical meristems
Located at root and shoot apex, responsible for primary growth (length increase).
- Lateral meristems (cambium)
Located laterally in older plant parts, parallel to organ axis, e.g., cork cambium (phellogen) and vascular cambium. Responsible for secondary growth (girth increase). Vascular cambium produces secondary vascular tissues; phellogen produces periderm replacing epidermis.
- Intercalary meristems
Found between permanent tissues, e.g., nodes of monocots and grass leaf bases. Allow length growth away from tips, useful if tips are damaged or eaten.
Types of growth in plants
- Primary growth – results in length increase, occurring in most monocots and herbaceous dicots, due to apical and sometimes intercalary meristems.
- Secondary growth
Occurs after primary growth due to lateral meristems, depositing new phloem and large amounts of secondary xylem (wood), increasing girth. Characteristic of trees and shrubs; some herbaceous plants show limited secondary thickening.
Primary growth
Primary growth in shoots
The shoot apex has four regions: cell division, cell elongation, cell differentiation, and permanent tissue. Cells become older moving away from apical meristems.
The region of cell division
The dome-shaped apical meristem has protoderm (forms epidermis), ground meristem (forms cortex and pith), and procambium (forms vascular tissues including pericycle, phloem, vascular cambium, and xylem).
Characteristics of apical meristems
- Small, cuboidal cells with thin cellulose walls and dense cytoplasm.
- Few small vacuoles.
- Tightly packed with no air spaces.
- Lack chloroplasts.
- Divide by mitosis; one daughter remains meristematic, others differentiate.
Zone of expansion or cell elongation
Daughter cells increase size mainly by osmotic water uptake. Length increase in stems and roots is mainly due to elongation here. Cell wall thickening by cellulose or lignin also occurs depending on cell type.
Zone of cell differentiation
Differentiation starts from procambium, producing protoxylem (inside) and protophloem (outside), parts of primary xylem and phloem. Cells between xylem and phloem retain division ability, forming vascular cambium.
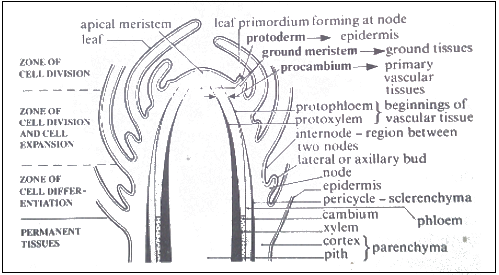
Formation of leaves and lateral buds
Leaves arise from small swellings or ridges of meristematic cells called primordia, appearing at nodes with internodes between. Nodes may be arranged in whorls, singly, or spirally. Primordia elongate rapidly, enclosing and protecting apical meristems physically and by heat from respiration, later growing into leaf blades.
The lateral bud (axillary bud)
Small groups of meristematic cells usually dormant but capable of division later, forming branches or specialized structures like flowers. They also form underground structures like rhizomes and tubers. Controlled by apical meristems, lateral buds develop in leaf and stem axils.
Primary growth in roots
Root growth regions:
- The root cap
Protects apical meristems as roots grow through soil. Made of parenchyma cells, constantly worn and replaced. Outer layer has mucilage to reduce friction and acts as gravity sensor.
- The zone of cell division
Includes:
- The quiescent centre
Tip of apical meristems, composed of initials with lower division rate than surrounding cells.
b) The protoderm, ground meristems, and procambium
Below quiescent centre, protoderm forms epidermis, ground meristems form cortex including endodermis, procambium forms primary phloem, vascular tissue, primary xylem, pith, and pericycle. Procambium in roots describes central cylinder.
- The zone of cell elongation
Cells elongate by osmotic water uptake into cytoplasm and vacuole, extending about 10 mm behind root tip, forcing root tip through soil.
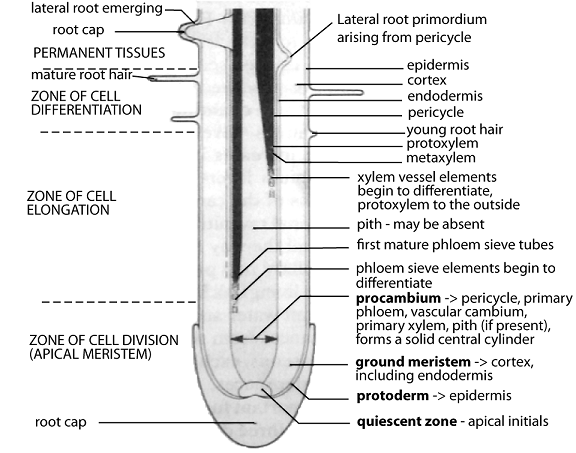
- The zone of cell differentiation
Cells specialize for functions. Phloem sieve elements differentiate from outside inward, becoming mature further from root tip. Xylem differentiates similarly (exarch), starting with protoxylem vessels transforming into metaxylem and mature xylem. Xylem spreads to root center, no pith develops. Root hairs develop from epidermis.
Formation of lateral roots, adventitious roots, and adventitious buds
Lateral roots: Arise from main root by pericycle cells resuming meristematic activity, forming new root apical meristem that grows through endodermis, cortex, and epidermis.
Adventitious roots and buds: Form in unusual positions by resumed meristematic activity. Adventitious roots develop independently of primary root, forming main rooting system in monocots from stem nodes. Important in propagation by stem cuttings and for clinging (e.g., ivy). Adventitious buds develop on roots, stems, or leaves, forming new branches.
- Adventitious roots – develop independently of primary root, important in propagation and clinging.
- Adventitious buds – develop on roots, stems, or leaves, forming new branches.
Secondary growth
Occurs after primary growth due to lateral meristems, increasing girth by depositing large amounts of xylem (wood). Two lateral meristems involved: vascular cambium (produces vascular tissue) and cork cambium (phellogen) (replaces ruptured epidermis).
The activity of the vascular cambium
Two cell types in vascular cambium:
a) Fusiform initials: narrow, elongated cells dividing by mitosis to form secondary phloem outside and secondary xylem inside. Xylem production exceeds phloem.
b) Ray initials: spherical cells dividing to form parenchyma, accumulating as rays between xylem and phloem.
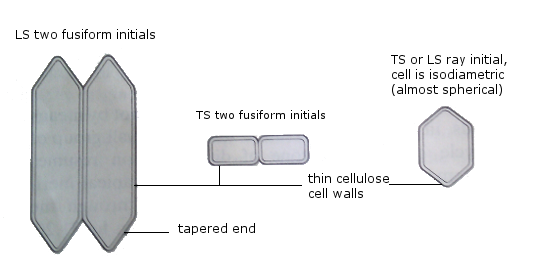
Secondary growth in woody dicotyledonous stem
Secondary growth or thickening in stem is due to deposition of large amounts of secondary xylem and lesser secondary phloem by fusiform initials of vascular cambium.
Vascular cambium is located between primary xylem and phloem of vascular bundles, called luto-fascicular cambium. Dicot vascular bundles form a ring. Initially, no cambium crosses the pith (medullary ray) between bundles. Cambium divides to form interfascicular cambium between bundles, completing the cambium ring.
The formation of secondary xylem and secondary phloem
The cambial layer is essentially one cell layer dividing parallel to epidermis. One daughter cell remains meristematic; the other differentiates into xylem (inside) or phloem (outside).
Continuous division adds cells to xylem and phloem, enlarging vascular bundles. As stem thickens, cambium circumference increases by radial division.
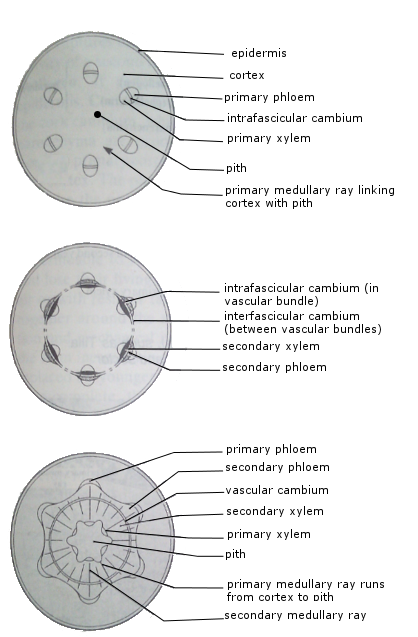
Formation of medullary rays
Medullary rays are parenchyma cells formed by ray initials, running from pith to cortex. The pith forms the stem center in dicots and roots of monocots. Extensions of pith as narrow parenchymatous strips are medullary or pith rays, primary from original ray initials and secondary from later initials.
Function of medullary rays
Rays maintain living connection between pith and cortex, facilitating rapid transport of water, minerals, and food substances across the stem.
Annual rings
Annual rings or growth rings are concentric layers of secondary xylem in perennial stems, each representing a seasonal increment or phase of xylem deposition.
In transverse sections, these appear as rings, commonly called annual rings because in temperate and some tropical woody plants, each ring corresponds to one year of growth.
Dendrochronology is the dating of wood by recognizing annual ring patterns.
Heartwood and sapwood
The heartwood
Central region of old trees where xylem ceases conducting and becomes blocked with dark deposits (tannins, gums, resins), making it hard and durable. It appears black due to these substances.
Sapwood
Outer region of old trees with recently formed xylem elements, lighter in color and containing some living cells. Performs physiological functions like water and nutrient conduction and food storage.
Cork and lenticels
Cork (phellogen)
Tissue formed by secondary lateral cambium (cork cambium) replacing ruptured epidermis due to stem circumference increase from secondary xylem growth.
Cork cells mature with walls impregnated by suberin, making them impermeable to water and gases. Cells die, lose contents, and fill with resins or tannins. Cork cells fit tightly to prevent desiccation, infection, and injury.
Lenticels are slit-like openings with loosely packed dead cells lacking suberin, produced by cork cambium, containing large air spaces for gaseous exchange between stem and environment. Without lenticels, gas exchange would be difficult as cork is impermeable.
Other cork cambium tissues: cork (phellem) forms outside, while one or two layers of parenchyma (phelloderm or secondary cortex) form inside, indistinguishable from primary cortex. Phellogen and phelloderm together form the periderm.
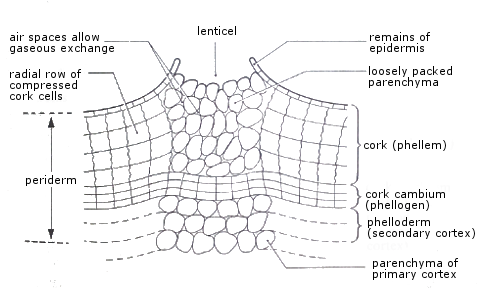
The bark
Bark refers to all tissues outside the vascular system or strictly those outside cork cambium in primary or secondary growth. It covers woody stems; peeling bark strips tissue down to vascular cambium. Bark consists of dead cells and cork layers.
Seed dormancy
Seed dormancy is a condition where seeds do not germinate despite favorable environmental conditions.
Causes of seed dormancy
1. Immaturity of the embryo
Newly harvested seeds need time for embryo maturation, undergoing internal changes called after-ripening. To end this dormancy, allow seeds sufficient time before sowing.
2. Hardness of the testa
Hard seed coat makes it impermeable to water and oxygen or physically strong to prevent embryo growth.
How to break this dormancy
- Physical damage or scarification of seed coat.
- Action of microorganisms like bacteria.
- Soaking for long periods and chemical action in soil.
Breaking of seed dormancy
Mechanism depends on dormancy type.
For primary dormancy
- Store seeds until embryo matures.
- Wet seeds with solvents like water and gibberellins to remove growth inhibitors.
- Remove testa by mechanical scarification or weakening chemicals (enzymes) to allow radicle and plumule emergence.
- Use organic solvents to dissolve waxy testa coating, making it permeable.
- Soak seeds in water to soften testa, allowing water and oxygen entry.
For secondary dormancy
- Provide appropriate temperature by chilling or temperature shock (boiling or hot water wash).
- Provide specific light intensity to raise gibberellin levels.
- Ensure sufficient moisture and oxygen.
Seed viability and germination
a) Seed viability
- Viability is the seed’s capacity to remain capable of germination under unfavorable conditions.
- A viable seed can germinate once dormancy causes are removed.
- Viability ensures germination when conditions and dormancy removal are met.
Factors governing seed viability
- Seed maturity
Immature seeds die and do not germinate due to incomplete embryo formation. Seeds need time to fully form embryos for active growth.
- Enzyme activity
- Germination depends on enzyme activation.
- Seeds with inactive enzymes cannot germinate and are inviable.
- Storage conditions
- Seed health
- Temperature: Optimum storage temperature preserves viability; high temperatures denature enzymes, preventing germination.
- Moisture: High moisture causes viability loss; dry storage is preferred.
- Aeration: Oxygen supply is needed for minimal metabolism; lack of aeration hinders viability.
Diseased seeds may lose viability due to fungal or bacterial infection.
- Storage duration
Viability varies by species; most annual seeds lose viability within a year. Some seeds, like Cassia bicapsularis and Cassia maltijuga, remain viable for over 100 years.
- Long storage may also lead to insect damage.




10 Comments